Développement visuoperceptif normal et pathologique
Un objectif de ce volet de recherche est de quantifier et de comparer la trajectoire du développement de différentes fonctions visuelles. Par exemple, nous évaluons le développement de différentes fonctions visuelles associées à différents niveaux de traitement cortical au sein de la hiérarchie du système visuel : de la sensibilité aux contrastes et à la perception du mouvement local, qui sont sous-tendus par les aires visuelles primaires, en passant par l'intégration des contours et la perception du mouvement global, qui impliquent des étapes de traitements additionnelles au sein des aires visuelles supérieures et extrastriées. Jusqu'à présent, les résultats suggèrent que les aspects de la perception visuelle qui nécessitent plusieurs étapes de traitement au sein du cortex visuel se développent plus lentement que celles qui requièrent moins d'étapes.
Projets récents :
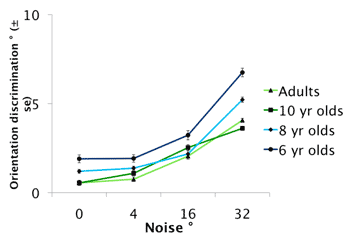
Developmental processes underlying orientation discrimination during childhood
Orientation-selectivity is critical for higher-level visual processes from edge extraction to the perception of natural images. Only a few studies documented the development of orientation discrimination during infancy, and developmental changes during childhood are practically unexplored. The goal of the present study was to chart the development of orientation discrimination during childhood as a function of contrast, spatial frequency, and orientation noise to understand the changes in its underlying mechanisms during development. Stimuli were circular two-dimensional luminance Gaussian noise with a diameter of 5 degrees. The noise was filtered in the Fourier domain with an anisotropic filter. Orientation discrimination thresholds were determined by means of a one-interval forced choice paradigm combined with an adaptive staircase. Participants had to decide whether the grating was tilted to the left or right of vertical. Six-year-olds are about 3.5 times less sensitive than adults at low (1 cpd) and mid (4 cpd) range spatial frequencies and they are 4.6 times less sensitive than adults at higher (12 cpd) spatial frequencies. Eight-year-olds are still less sensitive than adults by a 2-fold, and that equally so for each spatial frequency. Orientation discrimination is adult-like at 10 years of age for all conditions tested, falling within the range of 0.5 to 1.0 degree. Orientation discrimination is contrast invariant in children when tested with low (1 cpd) to mid range (4 cpd) spatial frequencies that have a contrast equal to or greater than 25%. However, thresholds increase by a factor of two for a spatial frequency of 12 cpd presented at a contrast of 25%. The introduction of noise orientation increases discrimination thresholds for each age group, but significantly more so for the 6- and 8-year-old children. Orientation discrimination matures slowly during childhood and it is influenced by spatial frequency and contrast in the developing visual system but not in the mature visual system. Because internal noise explains much of the functional immaturity in children, the main source of developmental variation in orientation discrimination most likely occurs at the initial stages of feature coding.
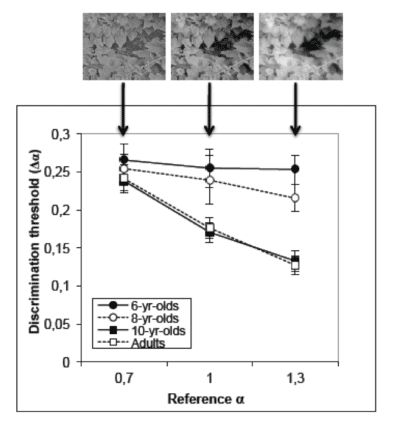
The developing visual system is not optimally sensitive to the spatial statistics of natural images
The adult visual system is optimally tuned to process the spatial properties of natural scenes, which is demonstrated by sensitivity to changes in the 1/f α amplitude spectrum. It is also well documented that different aspects of spatial vision, including those likely responsible for the perception of natural scenes (e.g., spatial frequency discrimination), do not become mature until late childhood. This led us to hypothesise that the developing visual system is not optimally tuned to process the spatial properties of real-world scenes. The present study investigated how sensitivity to the statistical properties of natural images changes during development. Thresholds for discriminating a change in the slope of the amplitude spectrum of a natural scene with a reference α of 0.7, 1.0, or 1.3 where measured in children aged 6, 8, and 10 years (n = 16 per age) and in adults (mean age = 23). Consistent with previous studies, adults were least sensitive for the shallowest α (i.e., 0.7) and most sensitive for the steepest α (i.e., 1.3). Six- and 8-year-olds had significantly higher discrimination thresholds compared to the 10-year-olds and adults for α's of 1.0 and 1.3, and 10-year-olds did not differ significantly from adults for any of the α's tested. These data suggest that sensitivity to detecting a change in the spatial characteristics of natural scenes during childhood may not be optimally tuned to the statistics of natural images until about 10 years of age. Rather, is seems that perception of natural images could be limited by the known immaturities in spatial vision (Ellemberg, Lepore & Turgeon, 2010). The question remains as to whether the adult's exquisite sensitivity to the spatial properties of the natural world is experience driven or whether it is part of our genetic programming that only fully expresses itself in late childhood.
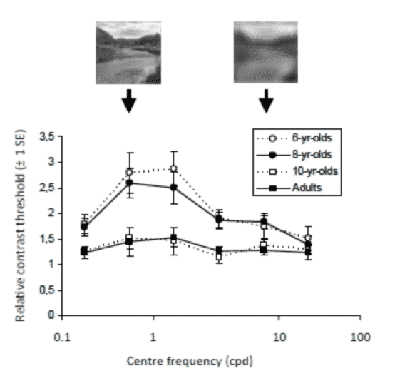
The development of natural image contrast sensitivity
The present study verified the hypothesis that the development of contrast sensitivity to the spatial frequency content of natural images extends beyond the development of contrast sensitivity when tested with luminance modulated sine-wave gratings. In the same group of children and adults, we compared contrast sensitivity with a series of natural images for which a specific slice of spatial content was removed (a narrow range of α values) and we also tested contrast sensitivity with a series of sine-wave gratings varying in spatial frequency. Thresholds were measured with a temporal forced-choice task combined with a QUEST staircase procedure. Our data show that for children and adults, detection thresholds for natural images were increased by the removal of spatial content and the shift in threshold was dependent on the spatial frequencies removed. Indeed, thresholds were most elevated when the frequencies removed were those that the visual system is most sensitive to. This suggests a close relationship between spatial frequency processing and natural image perception. However, unlike grating contrast sensitivity, which becomes adult-like at each frequency tested by about 7 years of age, contrast sensitivity for natural images only becomes adult-like by about 10 years of age. Further, sensitivity matures less rapidly for lower frequency filtered natural images than for the higher frequency filtered natural images. Together, these results indicate that the mechanisms involved in the processing of the spatial frequency content of natural images mature less rapidly than specific mechanisms tuned to a single spatial frequency and orientation. Finally, because of the important developmental differences between traditional contrast sensitivity compared to natural image contrast sensitivity, the latter might be a more ecologically relevant way assess the limits of visual perception in children and in individuals with a visual pathology.
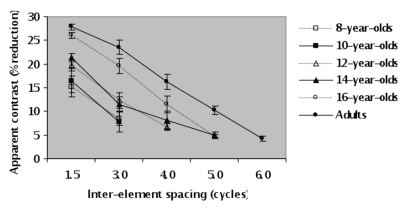
Spatial lateral interactions during childhood
We studied the development of spatial lateral interactions. Participants were adults (mean age = 23 years-old) and children aged 8, 10, 12, 14 and 16 years (n = 18 per age). We measured the apparent contrast of a foveally viewed Gabor as a function of the spacing of horizontally adjacent Gabors. Each Gabor had a contrast of 40%, a vertical carrier spatial frequency of 3 cycles per degree (γ = σ = 20 arc min), and a spread of 0.58 degrees. Inter-element spacing ranged from 1.5 to 6 cycles from the centre of the target to the centre of either of its adjacent flankers. The data make three points. First, at the smallest inter-element spacing, there is a reduction in the apparent contrast of the central Gabor for each age group tested. Second, at the smallest inter-element spacing, apparent contrast was significantly less reduced for the 8, 10, 12 and 14-year-olds compared to the two older groups. There was no significant difference between the 16-year-olds and the adults. Finally, compared to the adults, the measured reduction in apparent contrast extends over shorter distances for all other groups tested. Together, these results suggest that spatial lateral interactions mature very slowly and that they still have not reached adult-like characteristics by 16 years of age. This is consistent with recent findings that GABA expression in the human visual cortex does not become adult-like until late teens or even early adulthood [Boley et al., (2005). Society for Neuroscience].